Mặc dù trẻ hay thú sơ sinh mạnh khỏe đều có một hệ miễn dịch ‘có năng lực đầy đủ’ (competent) để thiết lập một đáp ứng miễn dịch đầy đủ đối với một kháng nguyên lạ hay vacxin. Tuy nhiên, về cơ chế hình thành đáp ứng miễn dịch thì có sự khác biệt rõ rệt giữa thú non và thú trưởng thành, vì ba yếu tố chính.

Thứ nhất, do sự phát triển của hệ thống miễn dịch, vì một số khía cạnh của đáp ứng miễn dịch bẩm sinh cũng như đáp ứng miễn dịch thích ứng chưa hoàn toàn phát triển khi mới sinh mà sẽ phát triển đầy đủ khi thú lớn dần.
Thứ hai, là do kinh nghiệm tiếp xúc với kháng nguyên lạ; cho đến khi sinh ra, mức độ tiếp xúc với kháng nguyên lạ của thú con là cực kỳ thấp.
Thứ ba, hệ thống miễn dịch của thú con trong thời kỳ bào thai bị ức chế tự nhiên nhằm giảm sản xuất các cytokine có tính kích viêm để tránh những phản ứng tiêu cực giữa thú mẹ và con trong thời kỳ mang thai.
Về mặt cơ thể học và sinh lý học, biểu mô đường tiêu hóa cũng như đường hô hấp ở thú sơ sinh đều có tính thấm cao hơn nhiều so với thú trưởng thành, chứng tỏ rằng sự toàn vẹn của các cơ quan chưa được thiết lập. Sự phân tiết các công cụ bảo vệ hóa học như các enzyme phân giải protein (protease) và các peptid kháng khuẩn cũng chưa nhiều . Thành phần hóa học và cấu trúc của lớp màng nhầy, hàng rào bắt giữ và ngăn chặn vi sinh vật xâm nhập, cũng chưa được phát triển
3.1. Miễn dịch bẩm sinh và miễn dịch qua trung gian tế bào
Về mặt miễn dịch học, so với cá thể trưởng thành, trẻ hay thú sơ sinh có số lượng bạch cầu có hạt (bạch cầu trung tính, bạch cầu ái toan, bạch cầu ái kiềm) thấp hơn. Ngoài ra, L-selectin là các phân tử bám dính của bạch cầu, có vai trò quan trọng trong sự di chuyển và thoát mạch của chúng (xem thêm trong phần ‘Bạch cầu thoát mạch’ và phần ‘Hệ thống miễn dịch bẩm sinh’). Bạch cầu có hạt ở thú sơ sinh lại biểu hiện ít phân tử L-selectin trên bề mặt, vì thế chúng ít có khả năng đáp ứng lại các tín hiệu hóa hướng động và thoát mạch kém (Hình).
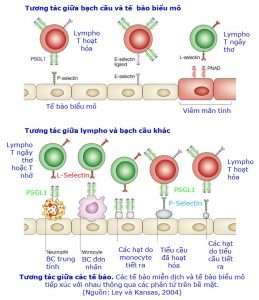
Hình ‘Tương tác giữa các tế bào’ cho thấy lympho T hoạt hóa dùng phân tử bề mặt PSGL1 hoặc E-selectin ligand để tiếp xúc với phân tử P-selectin hoặc E-selectin biểu hiện trên bề mặt tế bào biểu mô tại vùng viêm. Trong mô viêm mãn tính, thì ngoài P-selectin và E-selectin, tế bào biểu mô còn biểu hiện thêm phân tử bề mặt PNAD có khả năng tiếp xúc với phân tử L-selectin biểu hiện trên lympho T ngây thơ. Trong khi đó, khi các bạch cầu trung tính, bạch cầu đơn nhân hay các hạt do bạch cầu đơn nhân và tiểu cầu tiết ra đã tiếp xúc và chiếm đóng trên bề mặt tế bào biểu mô tại các mô viêm, thì chúng sẽ biểu hiện phân tử PSGL1 để tạo ra chỗ bám cho các lympho T ngây thơ, lympho T hoạt hóa hoặc lympho T nhớ.
Hơn nữa, các tế bào trình diện kháng nguyên (APC, gồm tế bào tua, đại thực bào, lympho B; xem thêm trong phần ‘Các tế bào trình diện kháng nguyên’) của thú non cũng hoạt động và phân tiết cytokine kém hơn. Lý do là vì chúng biểu hiện với số lượng thấp đối với các thụ thể và phân tử bề mặt quan trọng như MHC I, MHC II, thụ thể đồng hoạt hóa B7; đồng thời hoạt tính thực bào (phagocytosis), nhập bào (endocytosis) và hóa hướng động (chemotaxis) của chúng cũng chưa mạnh. Bên cạnh đó, hoạt động tầm soát và tiêu diệt tác nhân gây bệnh, tế bào bị nhiễm, tế bào biến thái của cở thể do tế bào NK phụ trách cũng còn yếu trên động vật sơ sinh. Để hiểu rõ thêm về chức năng của tế bào giết tự nhiên NK, xin tham khảo phần ‘Tế bào giết tự nhiên’.
Đáp ứng của lympho T với kháng nguyên ở động vật sơ sinh yếu hơn ở động vật trưởng thành, thể hiện thông qua mức sản xuất IFNγ thấp, đặc biệt là đáp ứng miễn dịch qua trung gian tế bào liên quan đến lympho CD4 TH1 và CD8 T (lympho CD8 T sau khi hoạt hóa còn được gọi là CTL hay Tc). Bởi vậy, nói chung là trên động vật sơ sinh, đáp ứng miễn dịch thích ứng đặc hiệu xảy ra mạnh hơn ở phần miễn dịch dịch thể do lympho B phụ trách với sự hỗ trợ của TH2. Mặc dù một số kháng nguyên vacxin mạnh có thể kích hoạt cả phần miễn dịch qua trung gian tế bào do lympho CD8 T và CD4 TDTH (lympho T quá mẫn muộn) phụ trách với sự hỗ trợ của CD4 TH1, nhưng hầu hết các vacxin trung bình đều chủ yếu kích thích miễn dịch dịch thể.
Để hiểu rõ hơn về hai phần của đáp ứng miễn dịch thích ứng đặc hiệu, xin tham khảo phần ‘Đáp ứng miễn dịch dịch thể’ và ‘Đáp ứng miễn dịch qua trung gian tế bào’. Hơn nữa, lượng lympho T có chức năng ức chế đáp ứng miễn dịch (gọi là Treg) lại hiện diện với số lượng lớn hơn trên động vật sơ sinh.
Tóm lại, các tế bào hay các chất tham gia đáp ứng miễn dịch bẩm sinh cũng như thích ứng đều còn đang ở trong trạng thái phát triển, chưa hoàn thiện trên động vật sơ sinh. Đặc trưng của các thành phần của hệ miễn dịch là chúng có mội quan hệ bổ trợ lẫn nhau. Vì vậy, người ta giải thích rằng đáp ứng yếu kém của hệ miễn dịch trên thú sơ sinh một phần chính là do thiếu vắng sự hỗ trợ lẫn nhau này.
3.2. Miễn dịch thể dịch
Số lượng lympho B trên động vật sơ sinh là rất cao, nhưng khả năng trưởng thành, biệt hóa và chuyển lớp chuỗi nặng của chúng còn hạn chế. Quá trình biệt hóa xảy ra là để biến đổi lympho B thành tương bào (plasma cell) chuyên sản xuất kháng thể, bao gồm một trong năm loại IgA, IgM, IgG, IgD và IgE. Ban đầu, tất cả mọi lympho B trưởng thành đều sản xuất IgM và IgD ở dạng gắn trên bề mặt (Hình ‘Quá trình trưởng thành và biệt hóa lympho B’, giai đoạn Lympho B trưởng thành). Sau đó, nhờ quá trình chuyển lớp chuỗi nặng, dưới sự kích thích của các cytokine và đặc biệt là cytokine của lympho TH2 mà chúng có thể biệt hóa thành các tương bào sản xuất các loại kháng thể dạng tiết (secreted immunoglobulin, sIg) gồm sIgA, sIgM, IgG và IgE.
Song song với quá trình biệt hóa thành tương bào, một số lympho B phát triển thành tập hợp các lympho B nhớ. Các lympho B nhớ này có khả năng nhận diện kháng nguyên đặc hiệu ở lần tiếp xúc tiếp theo và kích hoạt đáp ứng sản xuất kháng thể đặc hiệu một cách nhanh chóng và mạnh mẽ hơn so với lần tiếp xúc đầu tiên. Tập hợp lympho B nhớ bao gồm nhiều loại tế bào B nhớ khác nhau, mỗi loại mang trên bề mặt của nó duy nhất một loại kháng thể bề mặt (còn gọi là thụ thể BCR) khác nhau (gồm IgM, IgG, IgA, IgE) tương ứng với mỗi loại kháng thể đặc hiệu dạng tiết mà chúng sẽ sản xuất ra khi bị kích thích. Kháng thể IgD chỉ có ở dạng bám trên bề mặt tế bào lympho B ở giai đoạn lympho B trưởng thành, nó không được sản xuất ở dạng tiết và cho đến nay người ta vẫn chia hiểu biết nhiều về vai trò của IgD trong đáp ứng miễn dịch. Để hiểu rõ hơn về các loại kháng thể, xin tham khảo phần ‘Kháng thể’.

Trên động vật sơ sinh, ngoài kháng thể IgG nhận được từ mẹ thông qua nhau thai, qua sữa đầu và sữa, các kháng thể tự thân sản xuất ra rất thấp. IgA hầu như không có. IgM rất thấp mặc dù nó sẽ tăng nhanh chóng sau khi có sự kích thích của kháng nguyên đối với lympho B. Mức độ của IgE cũng rất thấp vì việc sản xuất IgE đòi hỏi phải có sự hỗ trợ của IL4 từ lympho TH2, mà lympho TH2 chưa hoạt động mạnh ở tuổi sơ sinh. Điều này một lần nữa cho thấy tầm quan trọng của việc cung cấp các men vi sinh sống qua đường tiêu hóa, nhất là trên động vật sơ sinh để chúng giúp kích thích phản ứng chuyển lớp chuỗi nặng và sản xuất IgA thông qua các yếu tố BAFF và APRIL được trình bày ở phần sau.
Bên cạnh đó, những quan sát khoa học hiện tại cho thấy rằng hệ vi sinh đường ruột giữ vai trò quan trọng trong việc điều hòa sự hình thành hệ thống phòng thủ của đường ruột:
1/ Chúng phát ra những tín hiệu hướng dẫn cho sự hình thành của các nhóm lympho bào then chốt;
2/ Chúng điều khiển sự chuyển lớp chuỗi nặng của lympho B;
3/ Chúng ức chế việc sản xuất thêm các lympho T điều hòa (Treg);
4/ Chúng kích thích sự hình thành nhóm lympho T giúp tiết interleukin 17 (gọi là TH17, tế bào này tiết cytokine như IL17, IL22 có vai trò kích thích tế bào biểu mô ruột sản xuất protein kháng khuẩn như nấm Candida, vi khuẩn Staphylococcus…, thiếu hụt TH17 thường mẫn cảm với các bệnh do vi khuẩn cơ hội);
5/ Chúng cũng gián tiếp quyết định sự hình thành của đáp ứng miễn dịch thích ứng đặc hiệu toàn thân thông qua việc điều hòa tỷ lệ lympho T giúp TH1 và TH2. Như đã đề cập ở trên, động vật sơ sinh có đáp ứng TH1 thấp, để cải thiện điều này, cần phải có sự hiện diện của nhiều loại vi sinh vật với số lượng cao trong đường ruột nhằm tạo ra các kích thích đối với hệ thống miễn dịch.
Cơ chế điều hòa: biểu mô ruột và các mô lympho của ruột có một số phương pháp điều hòa miễn dịch để tránh tình trạng xảy ra phản ứng quá mức dẫn đến sự hình thành những ổ viêm lớn gây tổn hại cơ quan. Người ta quan sát thấy rằng các tế bào trình diện kháng nguyên hiện diện trong mô ruột biểu hiện các thụ thể tầm soát vi sinh vật TLR ở mức rất thấp. Điều này giúp chúng tránh kích thích hình thành đáp ứng miễn dịch đối với những nhóm nhỏ vi khuẩn có lợi đường ruột mà thường xâm nhập xuyên qua biểu mô và lớp màng nhầy ruột.

Hình ‘Thụ thể TLR’ mô tả các loại thụ thể TLR chính đã được xác định hiện diện trên bề mặt tế bào. Chúng bao gồm TLR2, TLR4, TLR5 và TLR6. Trong đó, TLR4 và TLR5 hoạt động độc lập. TLR4 đã được chứng minh có khả năng nhận diện lipopolysaccharide (LPS) của các vi khuẩn gram âm, độc tố taxol ở thực vật, F protein của virus gây bệnh hô hấp RSV và một số protein của chính cơ thể vật chủ như HSP60 và fibronectin. TLR5 chuyên nhận diện kháng nguyên tiêm mao (Flagellin). Ngược lại, TLR2 và TLR6 luôn bắt cặp với nhau để hợp thành một loại thụ thể có khả năng nhận diện LPS của một số vi khuẩn như xoắn khuẩn Leptospira, vi khuẩn gram âm yếm khí Porphyromonas gingivalis, các lipoprotein, thành phần peptidoglycan của vi khuẩn, zymosan trong màng tế bào nấm men (yeast)…
Quá trình hoàn thiện của các cơ chế đề kháng và hệ thống miễn dịch sau sinh diễn ra nhanh hay chậm tùy thuộc vào nhiều yếu tố khác nhau. Trong đó, các yếu tố quan trọng nhất là sự cân bằng của hệ vi sinh đường ruột, thời gian và liều lượng mà những vi sinh vật cũng như những kháng nguyên mới lạ, lần đầu đi vào trong đường tiêu hóa qua thức ăn và nước uống. Những ngày đầu tiên đối với vật nuôi, hay những năm đầu tiên đối với con người là thời gian quan trọng nhất để hình thành đáp ứng miễn dịch và thói quen ăn uống. Những yếu tố khác như sữa mẹ, sữa thay thế hay việc sử dụng kháng sinh, men tiêu hóa cùng các sản phẩm khác trong thức ăn nước uống cũng góp phần ảnh hưởng đến sự cân bằng của hệ vi sinh đường ruột, tính toàn vẹn của biểu mô đường tiêu hóa và tính hoàn thiện của đáp ứng miễn dịch.
Biên soạn: TS. BSTY. Đinh Xuân Phát